|
GENETYKA
Genetyczne ubożenie rasy Azawakh
Autor:Elisabeth Naumann
Tłumaczenie na język polski: Judyta Kwaśniewska-Kaliszewicz
Prezentowany tekst to skrócona wersja tekstu prezentacji na temat rasy,
która została przedstawiona na spotkaniu poświęconym Azawakh'om DWZRV,
które odbyło się 25 stycznia 2003 roku.
Całą społeczność hodowców Azawakh'ów przeszło duże poruszenie w lecie 2002
roku. Niektórzy członkowie zgromadzenia francuskiego SLAG zasugerowali, że
dla polepszenia sytuacji rasy należałoby dopuścić do selektywnego
krzyżowania Azawakh'ów ze Sloughi. Pula genetyczna Azawakh'ów została
określona jako niewystarczająca w Europie. Czy te obawy są uzasadnione?
Jakie są ich powody? I jak temu zaradzić, (jeśli rzeczywiście jest to
konieczne)? Zanim odpowiem na powyższe pytania pozwolę sobie najpierw
wytłumaczyć, w najprostszy możliwy sposób, kilka podstawowych zagadnień
biogenetyki.
Komórka - podstawowy materiał
Komórka składa się z ciała komórki oraz z jądra, które niesie informację
genetyczną. Składa się ono z niteczek lub inaczej chromosomów, które
zawsze występują w parach. Pies posiada 78 chromosomów, czyli 39 par
chromosomów.
Materiał dziedziczny podzielony jest na dwie części. Nazywają się one
mikrosatelitami i są genetycznymi "kamieniami milowymi". Pies posiada 50
000 takich mikrosatelitów a każda z nich ma swoje specjalne miejsce na
chromosomie i jest ono różne u różnych psów. Im bardziej spokrewnione ze
sobą psy tym ich mikrosatelity są bardziej do siebie podobne.
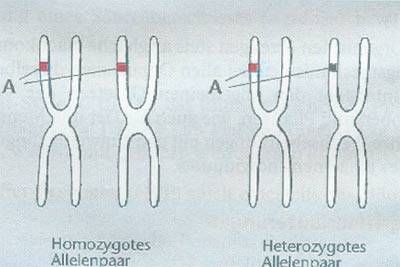
rys 1: homozygotyczna para allele
Geny - nośnik informacji genetycznej
Najmniejszą jednostką informacji genetycznej są geny. Chromosomy nie
istnieją indywidualnie, zawsze tworzą pary - homozygoty - każdy gen należy
do innego partnerskiego genu lub inaczej do homozygotycznego allele.
Zakładamy, że pies ma ponad 100 000 par allele, czyli 200 000 genów.
Miejsce, w którym allele są usytuowane na chromosomie nazywamy lokacją
genetyczną czyli locus. Specyficzny gen będzie można zawsze znaleźć w tej
samej lokacji na chromosomie. I ten konkretny gen będzie zawsze wpływał na
jedną konkretną cechę. Jeden chromosom w parze chromosomów pochodzi od
ojca a drugi od matki. Pies i suka dostarczają połowy genów.
Jeden allele może tłumić inny, co ma wpływ na wynik dziedziczenia. Jeśli
konkretny allele znajduje się na lokacji genu o mieszanej dziedziczności
to słabszy allele zostanie zdominowany a silniejszy, bardziej dominujący
zdeterminuje fenotyp. Te geny, które nie mogą się przebić nazywane są
genami recesywnymi.
Cechy recesywne mogą tylko być widoczne u potomków psów z mieszaną
dziedzicznością, jeśli ich nośnik w jednym ze zwierząt jest dziedzicznie
czysty. To znaczy, że te geny są usytuowane w parach w tej samej lokacji
genetycznej. Ten fakt odgrywa znaczącą rolę jeśli patrzymy na wady
genetyczne ponieważ wiele z nich jest dziedziczonych recesywnie. Jeśli
para allele zawiera tę samą informację genetyczną to pies będzie
homozygotyczny dla tej specyficznej cechy. Gdyby para allele była różna
dla danej cechy to pies był by dla tej cechy heterozygotyczny. Genetyczna
budowa psa jest w pewnych częściach czysto dziedziczona (homozygotycznie)
a w innych mieszanie dziedziczona (heterozygotycznie). Nie ma psów u
których wszystkie cechy byłyby w całości homozygotyczne.
Zgodnie z prawami genetyki
Jeśli spojrzymy na efekty dziedziczenia, możemy rozróżnić następujące
możliwości:
1.) jeden
gen wpływa na jedną cechę
2.) jeden gen wpływa na więcej niż jedną cechę
3.) większa ilość genów wpływa na całokształt cech, i w tym przypadku
mówimy o dziedziczności poligenetycznej, poligenetyce lub inaczej o
wieloczynnikowej genetyce
Zazwyczaj
to nie jeden specyficzny gen jest odpowiedzialny za konkretne cechy ale
jest to kombinacja większej ilości różnych genów. Taka poligenetyczna
dziedziczność jest podstawą prawie wszystkich cech charakterystycznych i
ilościowych. W rozumieniu cechy charakterystyczne mamy na myśli: wysokość,
wagę, kątowanie tylnych kończyn, kondycję i cechy naturalne.
Cechy jakościowe to szata i jej umaszczenie.
Geny mają jedynie wpływ na fenotyp w widoczny sposób (w wyglądzie psa)
jeżeli pojawi się ich określona minimalna ilość - jeśli przekroczą
określony próg. Takie graniczne (progowe) efekty są zauważalne gdy mamy do
czynienie z wadami genetycznymi. Niektóre wady to zmiany (mutacje)
zdrowych genów, które zostały przekazane recesywnie. Często pozostają w
ukryciu i nie ujawniają się przez całe pokolenia. Jeśli dwie mutacje się
połączą dopiero wtedy wada wyjdzie na "światło dzienne".
Tabela:
Stopień dziedziczenia dla różnych cech w procentach
(źródło: Hansen, Vererbung Am Hund, 2001)
|
Cecha
|
Dziedziczność
w %
|
|
Obszar rozmnażania
|
|
Płodność
|
10-15% nisko
|
|
Wielkość miotu
|
10-20% nisko
|
|
Jakość nasienia
|
15% nisko
|
|
Anatomia
|
|
Cechy anatomiczne
|
30-65% średnio do
wysoko
|
|
Wysokość w kłębie
|
40-65% wysoko
|
|
Długość ciała
|
40% średnio
|
|
Głębokość klatki
piersiowej
|
50% wysoko
|
|
Długość kłów
|
50% wysoko
|
Zachowanie
|
|
Instynkt łowiecki
|
10-30% nisko do
średnio
|
|
Temperament
|
30-50% średnio do
wysoko
|
|
Nerwowość
|
50% wysoko
|
|
Strach
|
45-60% wysoko
|
|
Wrażliwość na strzały
|
60-70% bardzo
wysoko
|
|
Instynkt tropienia
|
46% wysoko
|
|
Węch
|
39% średnio
|
|
Cechy obrończe
|
10% nisko
|
|
Połączone cechy charakteru
|
27-44% średnio
|
Dziedziczność to wpływ genotypu na fenotyp. Może się ona wahać pomiędzy 0
do 100%. Niemal żadna cecha nie jest dziedziczna w 100% (proszę porównać
do Claude Gaillard, "Der Zuchtwart" w "Rassehund, December 2002)
Selektywna hodowla jest sposobem na umocnienie lub stłumienie konkretnych
cech. Jedna z najprostszych i najoczywistszych zasad hodowli mówi, że
powinniśmy hodować tylko te osobniki, które łączą w sobie najlepsze cechy
rasy i są najbardziej zbliżone do standardu rasy. Oczywiście zawsze będzie
kwestia tego na ile sensowny jest standard rasy i kto powinien
determinować jej rozwój.

Ten pies rasy Azawakh w swojej ojczyźnie przedstawia typowe cechy
pożądanej budowy psa, takie jak: szeroki front, silne przednie kończyny,
obniżona linia grzbietu, mocna czaszka, silnie zarysowana dolna szczęka.
Te cechy ze względu na swój wysoki poziom dziedziczności mogą bardzo łatwo
zostać utracone poprzez częsty i wysoki inbred.

Czaszka Azawakh'a (Beliregion, Afryka Wschodnia), mocne zęby
i dobrze rozwinęta dolna szczęka są częścią "wyposażenia do przetrwania" w
ojczyźnie Azawakh'a.
Zazwyczaj hodowcy mają bogatą pulę nie spokrewnionych osobników, z których
mogą wybierać. Staje się to tym bardziej ważne gdy wady genetyczne
pokazują się przy wyborze homozygotycznym, co powoduje potrzebę ich
naprawy. Ta zasada może być stosowana w rasie Azawakh tylko do pewnego
stopnia z powodu mniejszej ilości osobników. Selektywna hodowla zawsze
oznacza zmiany genetyczne jak i straty genetyczne. Powtarzające się
selektywne kojarzenia zmieniają częstotliwość występowania konkretnych
genów w całej populacji hodowlanej.
Kiedy decydujemy o stratach genetycznych musimy przyjrzeć się dokładnie
rozmnażanemu zwierzęciu oraz skorzystać z pomocy współczynników inbredu
oraz współczynniku utraconych przodków. Ten pierwszy - współczynnik
inbredu - określa w jakim stopniu heterozygotyczność się zmniejsza na
korzyść homozygotyczności w konkretnym psie porównując do średniej w
rasie. Wzór liczący współczynnik inbredu (IK) wygląda następująco: IK-(1/2)n1+n2+1
Drugi ze współczynników to "utraceni przodkowie" (AVK) a wzór tego
współczynnika zaproponował prof. Schlegel z Uniwersytetu Wiedeńskiego.
Pokazuje on liczbę pojedynczych przodków (takich, którzy pojawili się
tylko raz w rodowodzie) pośród całej grupy przodków.
Przykład: w grupie przodków znanych do trzeciego pokolenia wstecz tylko
8miu z całej liczby 14stu przodków jest innych, wówczas współczynnik dla
całej grupy przodków będzie 8:14=0,55. I to oznacza utratę przodków w 55%.
AVK równy np. 60 oznacza utratę 40% heterozygotyczności.
Powracając do pytania od którego rozpoczęła się dyskusja "Ubożenie
genetyczne rasy Azawakh?" przeanalizowałam europejską populację rasy (900
osobników w latach 1999 do 2001) biorąc pod uwagę IK i AVK. Te wartości
zostały potem dodane do siebie a następnie podzielone przez ich ilość.
Wyniki pokazały stały spadek wartości AVK u populacji Azawakh'ów we
Francji. Pod koniec lat 90tych AVK spadł poniżej genetycznego progu 70ciu
%. To oczywiście z kolei działo się przy jednoczesnym wzroście
współczynnika inbredu ponad wartość krytyczną równą 20%. Powoduje to
poważny rozwój tzw. "wąskiego gardła" kiedy to mamy do czynienia z
sytuacją utraty początkowej szerokiej puli genetycznej Arawakh'ów z
importu z kolonialnej Afryki, czego nie da się naprawić z powodu
dramatycznego kurczenia się materiału genetycznego w dzisiejszej hodowli
francuskiej
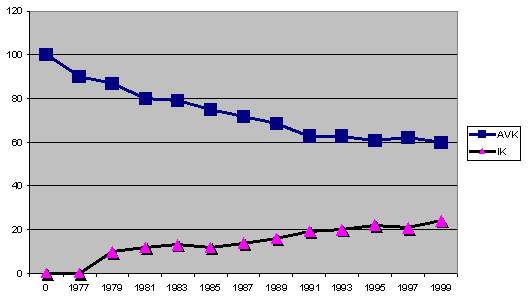
Graf 2: rozłożenie współczynników IK i AVK we francuskiej hodowli
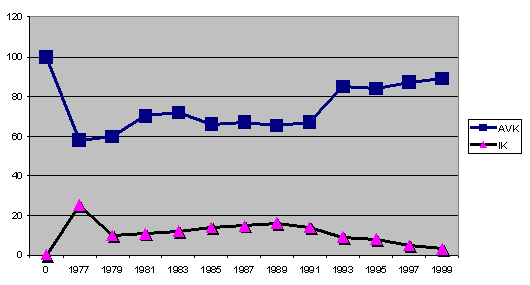
Graf 3: rozłożenie współczynników IK i AVK na przykładzie niemieckiej
hodowli
Niemiecka hodowla rozpoczęła się w okolicach lat 70tych XX wieku w bardzo
podobnych warunkach w jakich obecnie znajduje się populacja francuska.
Dzięki połączeniu w hodowli Azawakh'ów pochodzących z Francji z osobnikami
importowanymi z Afryki, niemiecka hodowla wydaje się być w znacznie
bezpieczniejszej sytuacji i od lat 90tych wzrasta. Oczywiście mogą istnieć
niezgodności i sprzeczności co do owej statystycznej średniej pomiędzy
różnymi liniami (graf 4-6).
Homozygotyczność jest uważana w normalnej hodowli zwierząt jako klucz do
optymalizacji rasy. Hodowla wsobna oznacza hodowanie psów homozygotycznych
- o homozygotycznym materiale genetycznym. Im większy współczynnik IK tym
większa możliwość, że potomstwo będzie homozygotyczne w pewnych genach.
Oczywiście otwartym pytaniem pozostaje, które z genów będą homozygotyczne
dzięki takiej hodowli - te pożądane czy te niepożądane geny. Postępowanie
w ten sposób w rasie, która już i tak jest uboga w pulę genetyczną i o
małym pogłowiu może doprowadzić do poważnych konsekwencji, głównie do
upadku całej rasy jako takiej. Geny recesywne w głównie poligenetycznym
dziedziczeniu najczęściej determinują wady.
Najmniej pożądane cechy, które występują w linii homozygotycznej są
błędami dziedziczenia.
Tylko potomstwo rozmnażanych jednostek, które niosą recesywnie
dziedziczony wadliwy homozygotyczny materiał, wykazują go w swoim
fenotypie. I to jest właśnie powodem, dla którego cechy, które poprzez
inbred powinny być wzmocnione, zostają sparowane ze wzmocnionymi wadami,
które są niepożądane. Lub w druga stronę - geny, które są niepożądane i
które staramy się wyeliminować poprzez inbred łączą się z genami, które są
najbardziej pożądane. W niewielkiej hodowli korzyści i ujemne rezultaty
takiego postępowania bardzo szybko się ujawniają.
Poprzez odbudowywanie na nowo populacji jakiejś rasy, krycia wsobne mogą
bardzo szybko dać się we znaki (często wymykając się spod kontroli) w
cechach zdefiniowanych przez hodowców lub kluby hodowlane. Jeśli dalsze
inbredy będą wiąż praktykowane to więcej coraz więcej ujemnych skutków
wyjdzie na jaw, taką sytuację może by określić jako depresję wsobną.< BR>
Negatywne wyniki inbredu nie pokazują się w ciągu jednej nocy, ani nie
pojawiają się w wszystkie na raz ani też w tym samym czasie.
Niektóre linie zdają się mieć wyższą tolerancję na inbred od innych, zanim
negatywne efekty się pokażą. Nawet w linii o bardzo bliskim inbredzie z
relatywnie wysokim IK i AVK, wady genetyczne mogą pozostać ukryte przez
dłuższy okres czasu. Dzięki tzw. szczęściu genetycznemu, mogą pozostać w
ograniczonym zakresie lub tylko ujawniać się w konkretnych cechach
psychicznych lub fizjologicznych, które nie są oczywiste dla hodowcy lub
właściciela a nawet mogą być tolerowane.
To mogłoby mieć dramatyczne skutki ponieważ te negatywne cechy w fenotypie
i w zachowaniu mogą mieć wpływ na sędziów jak i na ogólną opinię
wprowadzając w błąd i w przekonanie, że owe cechy są naturalne i w
zupełności pożądane. Częściowo dlatego właśnie, zasięg filozofii hodowli
jest zadziwiająco szeroki. Jako przykład , możemy porównać IK i AVK trzech
różnych miotów z trzech różnych hodowli Azawakh'ów.
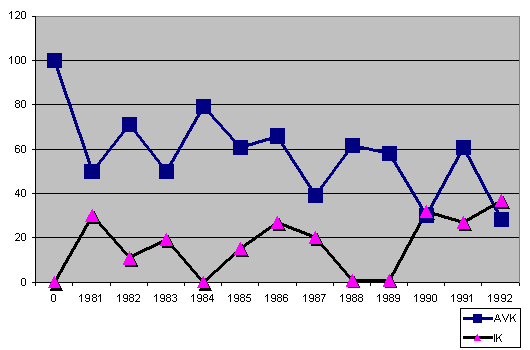
Graf 4: rozkład IK- i AVK na przkładzie hodowli wsobnej
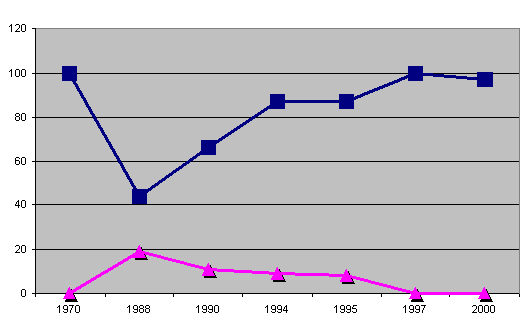
Graf 5: rozkład IK i AVK na przykładzie hodowli "outcrossing"
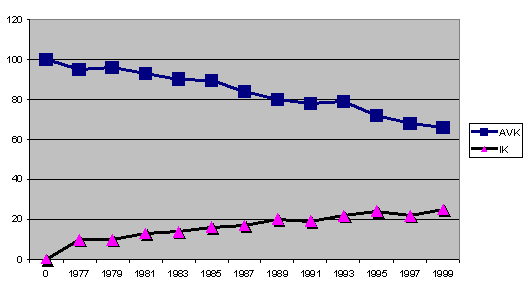
Graf 6: rozkład IK i AVK na przykładzie hodowli w obrębie jednej linii
Bliskie kojarzenie lub inbred nie muszą koniecznie prowadzić do klęski
puli genetycznej, zwłaszcza jeśli IK i AVK wrócą do równowagi poprzez
czasowe krzyżowanie nie spokrewnionych linii. Wartością progową dla IK
jest 10 a dla AVK uważa się 75. Bliskie kojarzenia - inbred - oznacza
zawsze, w każdym przypadku, straty nie do odzyskania i ryzyko
niepożądanych i powodujących choroby zmian.
Wydaje się, że francuski ZK miał właśnie te niebezpieczeństwa na myśli
(graf nr 4). W tym przypadku ich diagnoza byłaby obiektywnie właściwa, ale
ich propozycja zmiany tych trendów poprzez mieszanie Sloughi z
Aazawakh'ami po to by rozszerzyć pulę genetyczną, nie może być dobrym
sposobem. Prawdziwą alternatywą i szansą jest, praktykowane już przez
niemiecki ZK, wprowadzanie oryginalnego genetycznego dziedzictwa rasy
Azawakh.
|




{kind=link}
{kind=link}